


Санкт-Петербург, 2002 г.
РАЗДЕЛ 2. ТОКСИКОДИНАМИКА
ГЛАВА 2.1.
Механизмы токсического действия
Взаимодействие токсиканта или продуктов его превращения в организме со структурными элементами биосистем, лежащее в основе развивающегося токсического процесса, называется механизмом токсического действия. Взаимодействие осуществляется за счет физико-химических и химических реакции.
Токсический процесс, инициируемый физико-химическими реакциями, как правило, обусловлен растворением токсиканта в определенных средах (водной или липидной) клеток и тканей организма. При этом существенно изменяются физико-химические свойства среды-растворителя (рН, вязкость, электропроводность, сила межмолекулярных взаимодействий и т.д.). Особенность данного типа взаимодействия - отсутствие строгой зависимости качества развивающегося эффекта от химических свойств молекулы токсиканта. Таким образом, действуют на ткани все кислоты, щелочи, сильные окислители, некоторые органические растворители и лишенные специфической активности высокомолекулярные соединения.
Чаще в основе токсического действия лежат химические реакции токсиканта с определенным структурным элементом живой системы. Структурный компонент биологической системы, с которым вступает в химическое взаимодействие токсикант, называется его "рецептором" или "мишенью".
Механизмы токсического действия подавляющего большинства химических веществ в настоящее время неизвестны. В этой связи, очень многие описываемые ниже классы молекул и молекулярных комплексов, образующих организм, рассматриваются, по большей части, лишь как вероятные рецепторы (мишени) действия ядов. Рассмотрение их в этом ракурсе правомочно, поскольку в основе действия некоторых хорошо изученных токсикантов лежит взаимодействие с представителями именно этих классов биомолекул.
1. Определение понятия "рецептор" в токсикологии
Понятие "рецептор" весьма емкое. Наиболее часто в биологии его используют в следующих смыслах:
1. Общее понятие. Рецепторы - это участки относительно специфического связывания на биосубстрате ксенобиотиков (или эндогенных молекул), при условии, что процесс связывания подчиняется закону действующих масс. В качестве рецепторов могут выступать целые молекулы белков, нуклеиновых кислот, полисахаридов, липидов или их фрагменты. В отношении фрагмента биомолекулы, которая непосредственно участвует в образовании комплекса с химическим веществом, часто используют термин - "рецепторная область". Например, рецептором оксида углерода в организме является молекула гемоглобина, а рецепторной областью - ион двухвалентного железа, заключенный в порфириновое кольцо гема.
2. Селективные рецепторы. По мере эволюционного усложнения организмов формируются специальные молекулярные комплексы - элементы биологических систем, обладающие высоким сродством к отдельным химическим веществам, выполняющим функции биорегуляторов (гормоны, нейромедиаторы и т.д.). Участки биологических систем, обладающие наивысшим сродством к отдельным специальным биорегуляторам, получили название "селективные рецепторы". Вещества, взаимодействующие с селективными рецепторами в соответствии с законом действующих масс, называются лигандами селективных рецепторов. Взаимодействие эндогенных лигандов с селективными рецепторами имеет особое значение для поддержания гомеостаза.
Многие селективные рецепторы состоят из нескольких субъединиц, из которых лишь часть имеет участки связывания лигандов. Нередко термин "рецептор" используют для обозначения только таких лиганд-связывающих субъединиц.
3. Постоянные рецепторы - это селективные рецепторы, строение и свойства которых кодируется с помощью специальных генов или постоянных генных комплексов. На уровне фенотипа изменение рецептора путем генной рекомбинации развивается чрезвычайно редко. Возникающие порой в ходе эволюции вследствие полигенетических трансформаций изменения аминокислотного состава белка, формирующего селективный рецептор, как правило, слабо сказывается на функциональных характеристиках последнего, его сродстве к эндогенным лигандам и ксенобиотикам.
К числу постоянных рецепторов относятся:
- рецепторы нейромедиаторов и гормонов. Как и другие селективные рецепторы, эти рецепторы способны избирательно взаимодействовать и с некоторыми ксенобиотиками (лекарствами, токсикантами). Ксенобиотики могут при этом выступать как в качестве агонистов, так и антагонистов эндогенных лигандов. В итоге активируется или подавляется некая биологическая функция, находящаяся под контролем данного рецепторного аппарата;
- энзимы - белковые структуры, селективно взаимодействующие с субстратами, превращение которых они катализируют. Энзимы также могут взаимодействовать с чужеродными веществами, которые в этом случае становятся либо ингибиторами, либо аллостерическими регуляторами их активности;
- транспортные протеины - избирательно связывают эндогенные лиганды определенного строения, осуществляя их депонирование или перенос через различные биологические барьеры. Токсиканты, взаимодействующие с транспортными протеинами, также выступают либо в качестве их ингибиторов, либо аллостерических регуляторов.
4. Рецепторы с изменяющейся структурой. В основном это антитела и антигенсвязывающие рецепторы Т-лимфоцитов. Рецепторы данного типа формируются в клетках предшественниках зрелых клеточных форм вследствие индуцированной внешними воздействиями рекомбинации 2 - 5 генов, контролирующих их синтез. Если рекомбинация осуществилась в процессе дифференциации клеток, то в зрелых элементах будут синтезироваться рецепторы только определенного строения. Таким способом формируются селективные рецепторы к конкретным лигандам, а пролиферация приводит к появлению целого клона клеток, содержащих эти рецепторы.
Как следует из приведенных определений, в биологии термин "рецептор" в основном используется для обозначения структур, принимающих непосредственное участие в восприятии и передаче биологических сигналов, и способных избирательно связывать помимо эндогенных лигандов (нейромедиаторов, гормонов, субстратов) некоторые чужеродные соединения.
В токсикологии (как и фармакологии) термином "рецептор" обозначают любой структурный элемент живой (биологической) системы, с которым вступает в химическое взаимодействие токсикант (лекарство). В таком прочтении это понятие ввел в химеобиологию в начале ХХ века Пауль Эрлих (1913).
Спектр энергетических характеристик рецептор-лигандного взаимодействия необыкновенно широк: от формирования слабых, легко разрушающихся связей, до образования необратимых комплексов (см. выше). Характер взаимодействия и структура сформировавшегося комплекса зависят не только от строения токсиканта, конформации рецептора, но и от свойств среды: рН, ионной силы и т.д. В соответствии с законом действующих масс, количество образовавшихся комплексов вещество-рецептор определяется энергией взаимодействия (сродством) и содержанием обоих компонентов реакции (вещества и рецептора к нему) в биологической системе.
Рецепторы могут быть "немыми" и активными. "Немой" рецептор - структурный компонент биологической системы, взаимодействие которого с веществом не приводит к формированию ответной реакции (например, связывание мышьяка белками, входящими в состав волос, ногтей). Активный рецептор - структурный компонент биологической системы, взаимодействие которого с токсикантом инициирует токсический процесс. Для того, чтобы избежать терминологических трудностей, для обозначения структурных элементов, взаимодействуя с которыми токсикант инициирует токсический процесс, вместо термина "рецептор", часто используют термин "структура-мишень".
Принимаются постулаты:
- токсическое действие вещества выражено тем сильнее, чем большее количество активных рецепторов (структур-мишеней) вступило во взаимодействие с токсикантом;
- токсичность вещества тем выше, чем меньшее его количество связывается с "немыми" рецепторами, чем эффективнее оно действует на активный рецептор (структуру-мишень), чем большее значение имеет рецептор и повреждаемая биологическая система для поддержания гомеостаза целостного организма.
Любая клетка, ткань, орган содержат огромное количество потенциальных рецепторов различных типов ("запускающих" различные биологические реакции), с которыми могут вступить во взаимодействие лиганды. С учетом вышесказанного, связывание лиганда (как эндогенного вещества, так и ксенобиотика) на рецепторе данного типа является избирательным лишь в определенном диапазоне концентраций. Увеличение концентрации лиганда в биосистеме приводит к расширению спектра типов рецепторов, с которыми он вступает во взаимодействие, а следовательно, изменению его биологической активности. Это также одно из фундаментальных положений токсикологии, доказанное многочисленными наблюдениями.
Мишенями (рецепторами) для токсического воздействия могут быть:
- структурные элементы межклеточного пространства;
- структурные элементы клеток организма;
- структурные элементы систем регуляции клеточной активности.
2. Действие токсиканта на элементы межклеточного пространства
Каждая клетка организма окружена водной средой - интерстициальной или межклеточной жидкостью. Для клеток крови межклеточной жидкостью является плазма крови. Основные свойства межклеточной жидкости: её электролитный состав и определенное осмотическое давление. Электролитный состав определяется главным образом содержанием ионов Na+, K+, Са2+, Cl-, HCO3- и др.; осмотическое давление - присутствием белков, других анионов и катионов. Межклеточная жидкость содержит многочисленные субстраты для клеточного обмена, продукты метаболизма клеток, молекулы-регуляторы клеточной активности.
Попав в межклеточную жидкость, токсикант может изменять её физико-химические свойства, вступать в химическое взаимодействие с её структурными элементами. Изменение свойств межклеточной жидкости немедленно приводит к реакции со стороны клеток. Возможны следующие механизмы токсического действия, обусловленные взаимодействием токсиканта с компонентами межклеточной жидкости:
1. Электролитные эффекты. Нарушение электролитного состава наблюдается при отравлении веществами, способными связывать ионы. Так, при интоксикациях фторидами (F-), некоторыми комплексообразователями (Na2ЭДТА, ДТПА и др.), другими токсикантами (этиленгликолем, метаболизируящим с образованием щавелевой кислоты), происходит связывание ионов кальция в крови и межклеточной жидкости, развивается острая гипокальциемия, сопровождающаяся нарушениями нервной деятельности, мышечного тонуса, свертывающей системы крови и т.д. Нарушение ионного баланса, в ряде случаев, может быть устранено введением в организм растворов электролитов.
2. рН-эффекты. Интоксикация рядом веществ, не смотря на высокую буферную емкость межклеточной жидкости, может сопровождаться существенным нарушением кислотно-основных свойств внутренней среды организма. Так, отравление метанолом приводит к накоплению в организме муравьиной кислоты, вызывающей тяжелый ацидоз. Изменение рН интерстициальной жидкости может быть также следствием вторичных токсических эффектов и развиваться вследствие нарушения процессов биоэнергетики, гемодинамики (метаболический ацидоз/алкалоз), внешнего дыхания (газовый ацидоз/алкалоз). В тяжелых случаях нормализовать рН можно, вводя пострадавшему буферные растворы.
3. Связывание и инактивация структурных элементов межклеточной жидкости и плазмы крови. В плазме крови содержатся структурные элементы, обладающие высокой биологической активностью, способные стать мишенью действия токсикантов. К их числу относятся, например, факторы свертывающей системы крови, гидролитические ферменты (эстеразы), разрушающие ксенобиотики и т.д. Следствием такого действия может стать не только интоксикация, но и аллобиоз. Например, угнетение активности три-о-крезилфосфатом (ТОКФ) карбоксилэстераз плазмы крови, разрушающих фосфорорганические соединеня (ФОС), приводит к существенному повышению токсичности последних.
4. Нарушение осмотического давления. Существенные нарушения осмотического давления крови и интерстициальной жидкости при интоксикациях, как правило, носят вторичный характер (нарушение функций печени, почек, токсический отек легких). Развивающийся эффект пагубным образом сказывается на функциональном состоянии клеток, органов и тканей всего организма.
3. Действие токсикантов на структурные элементы клеток
Структурными элементами клеток, с которыми взаимодействуют токсиканты, как правило, являются:
- белки;
- нуклеиновые кислоты;
- липидные элементы биомембран;
- селективные рецепторы эндогенных биорегуляторов (гормонов, нейромедиаторов и т.д.).
3.1. Взаимодействие токсикантов с белками.
Основные функции белков: транспортная, структурная, энзиматическая (белки - биологические катализаторы). Токсический эффект может развиваться при нарушении каждой из этих функций.
Нарушение свойств белков химическим веществом возможно различными способами, зависящими как от структуры токсиканта, так и от строения и функций белка. Возможны: денатурация белка, блокада его активных центров, связывание активаторов и молекул, стабилизирующих протеин, и т.д.
К числу веществ, денатурирующих белки, относятся крепкие щелочи, кислоты, окислители, ионы тяжелых металлов. В основе денатурации лежит повреждение внутрибелковых связей, поддерживающих вторичную, третичную структуру протеина. При этом наиболее часто токсиканты взаимодействуют с СООН-, NH-, OH-, SH-группами аминокислот, образующих белки. Многочисленные токсиканты, связывающиеся с SH-группами, называются тиоловыми ядами. К числу тиоловых ядов прежде всего следует отнести тяжелые металлы, такие как ртуть, мышьяк, сурьма, таллий, органические соединения этих металлов (метилртуть, люизит и т.д.). Другие металлы более активно взаимодействуют с карбоксильными группами (свинец, кадмий, никель, медь, марганец, кобальт).
Особое значение в токсикологии придают действию ксенобиотиков на энзимы.
3.1.1. Энзимы
Роль энзимов в обеспечении процессов жизнедеятельности огромна. Неудивительно, что вещества, модулирующие их активность, обладают высокой биологической активностью, порой являются высокотоксичными веществами.
Энзимы - это белки, выполняющие функции биологических катализаторов. Они ускоряют достижение состояния равновесия обратимых экзергических реакций, типа: АВ
Скорость каталитических превращений веществ в организме определяется специфической активностью энзимов, их содержанием в клетках и тканях, наличием субстратов и регуляторов активности в среде. При нормальных условиях интенсивность процессов поддерживается на определенном уровне. Регуляция осуществляется различными механизмами, среди которых основные - модуляция количества энзимов, их специфической активности, изменение биодоступности субстратов и т.д. Основные пути регуляции ферментативных процессов представлены на рисунке 1.
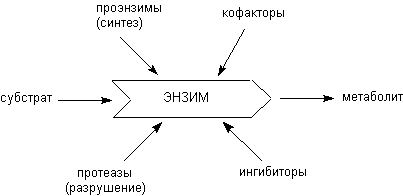
Рисунок 1. Пути регуляции ферментативных процессов
Возможные механизмы модуляции активности энзимов химическими веществами представлены в таблице 1.
Таблица 1. Механизмы изменения каталитической активности энзимов в условиях действия токсикантов
Усиление каталитической активности - усиление синтеза энзимов - блокада разрушения энзимов - активация энзимов |
Угнетение каталитической активности - угнетение синтеза энзимов - ускорение разрушения энзимов - угнетение специфической активности |
Изменение конформации энзимов |
3.1.1.1. Усиление каталитической активности
Усиление синтеза энзимов может быть вызвано поступлением в организм токсикантов-индукторов.
Физиологическими индукторами синтеза энзимов являются многие субстраты и вещества, повышающие содержание коэнзимов в биосредах. Некоторые гормоны выступают в качестве индукторов синтеза белка. Так, трииодтиронин у тиреоэктомированных крыс существенно увеличивает содержание глюкозо-6-фосфатазы и НАДН-цитохром-с-редуктазы в микросомах печени. Стероидные гормоны - активные индукторы синтеза ферментов, например, триптофанпирролазы и др.
Особое значение для токсикологии имеет явление индукции энзимов, участвующих в метаболизме ксенобиотиков.
К числу индукторов относятся барбитураты, циклические углеводороды, полигалогенированные полициклические углеводороды и многие другие. Токсичность такого известного токсиканта, как 2,3,7,8-тетрахлорпарадибензодиоксин (диоксин, ТХДД) в настоящее время связывают именно со способностью вызывать индукцию синтеза ферментов. Среди индукторов многие - канцерогены. Например, 3,4-бенз(а)пирен, 5-метилхолантрен.
Активность энзима зависит от наличия в биосредах кофакторов или простетических групп. Функции кофакторов выполняют различные витамины и ионы металлов. Их поступление в организм необходимо, однако передозировка сопровождается интоксикацией. Особенно опасно перенасыщение организма жирорастворимыми витаминами (А, Д). Стойкое повышение содержания ионов кальция в цитоплазме клеток, отмечаемое при интоксикациях некоторыми веществами, сопровождается чрезмерной активацией ряда ферментов (см. ниже).
Существенное влияние на активность ферментов оказывают вещества, блокирующие процессы их разрушения. Все белковые молекулы в организме имеют определенное время жизни. Процессы непрерывающегося синтеза уравновешиваются столь же постоянным разрушением белка. Период полусуществования молекул энзимов колеблется в широких пределах. Например, для альдолазы мышечной ткани крыс он составляет около 20 дней, для каталазы - 1 день, для триптофанпирролазы печени - 2 часа. В процессе разрушения энзимов принимают участие протеазы и эндопептидазы. Разрушение короткоживущих белков осуществляется также энзимами аппарата Гольджи. Ингибиторами разрушения энзимов (и других белков) являются ингибиторы протеаз/пептидаз. К их числу, относятся некоторые карбамилфосфаты и др.
Разрушение SH-содержащих энзимов иногда начинается с окисления этих групп. Ксенобиотики с высоким восстановительным потенциалом, защищая сульфгидрильные группы, могут предотвращать разрушение энзимов. Эти эффекты также могут лежать в основе токсического процесса.
Особую роль в токсикологии играют механизмы активации лизосомальных ферментов, вызывающих, при выходе в цитоплазму, аутолиз клеток. Посредством такого механизма действуют на организм многочисленные вещества, например, иприты, СCl4, и т.д.
3.1.1.2. Угнетение каталитической активности
Снижение активности энзимов при действии токсикантов может быть следствием трех эффектов: подавления процессов синтеза апофермента и кофакторов, активации разрушения, угнетения специфической активности.
К числу наиболее распространенных кофакторов, помимо металлов, относятся железопорфирины, флавины, никотинамид-адениндинуклеотид (НАД), пиридоксальфосфат, тиаминпирофосфат и др. Отчасти эти вещества синтезируются в организме животных и человека, отчасти попадают с пищей в форме витаминов. Некоторые вещества являются конкурентами кофакторов энзимов. Так, дикумарол конкурентно препятствует утилизации печенью витамина К, необходимого для синтеза протромбина, поэтому через 24 - 96 ч после поступления вещества в организм в токсических дозах возможно развитие кровотечений угрожающих жизни.
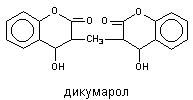
Некоторые токсиканты нарушают образование коферментов, предшественники которых поступают в организм с пищей. Так, гидразин и его производные, взаимодействуя с пиридоксалем, содержащимся в клетках, образуют пиридоксальгидразоны (рисунок 2), которые, в свою очередь, угнетают активность пиридоксалькиназы и блокируют тем самым синтез в организме пиридоксальфосфата. В итоге понижается активность большого числа энзимов, кофактором которых является пиридоксальфосфат (декарбоксилазы, трансаминазы и т.д.).
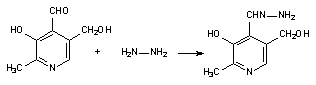
Рисунок 2. Взаимодействие гидразина с пиридоксалем с образованием пиродоксальгидразона
К числу полностью синтезируемых в организме кофакторов относятся железопорфирины. Блокада их синтеза приводит к тяжелым последствиям. Так, хроническое отравление свинцом сопровождается нарушением синтеза гема, вследствие чего развивается дефицит гемопротеинов (гемоглобина, миоглобина, гем-содержащих энзимов).
Активация процесса разрушения энзимов токсикантами, как механизм их токсического действия, встречается редко. Катаболизм некоторых энзимов усиливается на фоне хронической интоксикации стероидными препаратами и их аналогами.
Наиболее часто в основе интоксикации лежит угнетение токсикантом специфической активности энзимов. Выделяют следующие механизмы ингибиторного действия ксенобиотиков:
1. Конкурентное ингибирование. В основе взаимодействия лежит конкуренция токсиканта с субстратом за активный центр энзима. При этом реализуются две возможности:
а) токсикант вступает в превращение вместо субстрата (конкурентные субстраты). Так, некоторые эфиры холина (пропионилхолин, бутирилхолин и др.) гидролизуются ацетилхолинэстеразой вместо ацетилхолина. Конкурентным ингибитором аконитазы, одного из энзимов цикла трикарбоновых кислот, участвующих в превращении лимонной кислоты в аконитовую, является фторлимонная кислота, образующаяся в процессе метаболических превращений опасного токсиканта фторуксусной кислоты (рисунок 3);
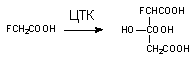
Рисунок 3. Превращение фторуксусной кислоты во фторлимонную в процессе метаболизма в цикле трикарбоновых кислот (ЦТК)
б) взаимодействие токсиканта с активным центром не приводит к его метаболизму (стабильные ингибиторы). Примерами таких токсикантов являются карбаматы - ингибиторы холинэстеразы.
Конкурентный тип ингибирования развивается также при образовании прочных ковалентных связей между токсикантами и активными центрами некоторых энзимов. Этот вид ингибирования приводит к полному прекращению ферментативной активности. Таким способом фосфорорганические соединения взаимодействуют с ацетилхолинэстеразой.
Часто, конкурентные ингибиторы энзимов структурно напоминают их субстраты.
2. Неконкурентное ингибирование. В данном случае токсикант взаимодействует с добавочным, аллостерическим, центром энзима, изменяя при этом конформацию активного центра и снижая, тем самым, его сродство к субстрату. Таким способом упомянутая выше фторлимонная кислота угнетает активность транслоказы, фермента, обеспечивающего активный транспорт цитрата через мембраны митохондрий, а мышьяк и его соединения - SH-содержащие энзимы.
3. Прочие. Для осуществления ферментативной активности энзимы нуждаются в присутствии в среде ионов металлов: Mg2+, Ca2+, K+, Mn2+, Zn2+, Co2+ и др. Связывание этих металлов токсикантами приводит к угнетению активности. Таков механизм токсического действия комплексообразователей (ЭДТА, ДТПА, дитизона и др.), салициловой кислоты и др. Особое токсикологическое значение имеют вещества, взаимодействующие с железом, кобальтом, медью, входящими в структуру более сложных простетических групп энзимов (гем-содержащие энзимы, цитохромы, каталаза, пероксидаза, гемоглобин, миоглобин). К числу подобных токсикантов относятся цианиды, сульфиды, азиды, монооксид углерода и др.
Некоторые энзимы находятся под постоянным контролем специальных кооперационных систем. Так, система GSH/GSSG (восстановленный/окисленный глутатион) регулирует активность SH-энзимов (пирофосфатаза, фосфоглицеральдегид-дегидрогеназа, гемоглобин-редуктаза и др.). Токсиканты, понижающие содержание восстановленного глютатиона в тканях, такие как гидроксиламин, фенилгидразин, дихлорэтан и др., подавляют активность этих ферментов.
Одна из патохимических классификаций преимущественно ферментных ядов разработана А.А. Покровским (1962) (таблица 2).
Таблица 2. Патохимическая классификация ядов
МЕХАНИЗМ ДЕЙСТВИЯ ЯДОВ НА ФЕРМЕНТЫ |
ХАРАКТЕРНЫЕ ПРЕДСТАВИТЕЛИ ТОКСИЧНЫХ ВЕЩЕСТВ |
Структурные аналоги данного субстрата, взаимодействующие с ним по типу "конкурентного торможения" |
Фосфорорганические соединения и другие антихолинэстеразные соединения, Циклосерины, Галоидопроизводные пиримидина и др. |
Предшественники структурных аналогов субстрата, из которых образуются ингибиторы ферментов в процессе "летального синтеза" |
Фторацетат, хлорацетат Галоидсодержащие производные пиримидина и пурина, Метиловый спирт, Этиленгликоль и др. |
Структурные аналоги коферментов, антивитамины. Конкурентное торможение |
Антивитамины РР, В6 и др. |
Соединения, тормозящие биосинтез ферментов. Крнкурентное торможение; в отдельных случаях неконкурентное |
Структурные аналоги природных аминокислот (фторфенилаланин, азатриптофан и др.), Некоторые антибиотики: пенициллин, левомицетин, ауреомицин и др. |
Соединения, блокирующие функциональные группы белка или кофермента. Неконкурентное торможение |
Цианиды, Сероводород, Окись углерода, Метгемоглобинообразователи, Соединения, связывающие SH-группы и др. |
Соединения, нарушающие связи атомов металлов в молекуле фермента |
Хелатирующие соединения: ЭДТА, ДТПА, 8-оксихинолидин и т.п. |
Соединения, разобщающие сочетанную деятельность ферментов |
Разобщители клеточного дыхания и окислительного фосфорилирования (динитрофенол), Фториды, Некоторые наркотики и др. |
Соединения, денатурирующие белок (грубое нарушение структуры белковой молекулы) |
Крепкие кислоты, щелочи, тяжелые металлы, органические растворители и др. |
Биологические яды, содержащие ферменты, разрушающие структурные элементы клеток и тканей, образующие в организме токсичные вещества |
Яды змей и насекомых, Бактериальные токсины |
Аналоги медиаторов |
Ингибиторы моноаминоксидазы и др. |
3.1.1.3. Биологические последствия действия токсикантов на энзимы
Поскольку все процессы в живых организмах протекают при участии энзимов, и все фундаментальные свойства живых систем неразрывно связаны с нормальным течением этих процессов, теоретически любые проявления жизни могут быть нарушены теми или иными токсикантами, изменяющими активность энзимов.
3.2. Взаимодействие токсикантов с нуклеиновыми кислотами.
Дезоксирибонуклеиновые кислоты - основной компонент хромосомного аппарата клеток. Рибонуклеиновые кислоты представлены информационной, транспортной, рибосомальной РНК. Их функция - участие в синтезе белка. Многие ксенобиотики вступают во взаимодействие с нуклеиновыми кислотами, изменяя их свойства.
1. Химическая модификация нуклеиновых кислот.
К числу веществ, вступающих в химическое взаимодействие с нуклеиновыми кислотами, относятся нитриты, сернистый, азотистый, кислородный иприты, этиленоксид, этиленимин, гидразин и его производные, гидроксиламин, нитрозамины, аренокисды, полициклические углеводороды, метаболиты афлатоксинов, соединения мышьяка и многие другие вещества. Эти токсиканты, образуют ковалентные связи с аминогруппами пуриновых и пиримидиновых оснований, входящих в структуру нуклеиновых кислот (рисунок 4). Измененные таким образом молекулы ДНК могут подвергаться дальнейшей ферментативной и неферментативной трансформации вплоть до разрушения под воздействием эндонуклеаз.
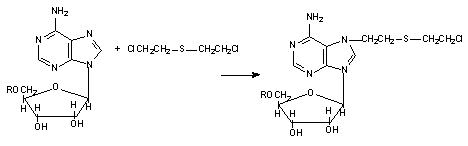
Рисунок 4. Взаимодействие аденозина с ипритом
Вещества с бифункциональными активными группами (иприты) могут образовывать с двунитевой молекулой ДНК перекрестные связи, при этом становиться невозможным расхождение нитей "двойной спирали", необходимое для обеспечения синтеза белков, клеточного деления.
Токсиканты способны вступать во взаимодействие не только с пуриновыми и пиримидиновыми основаниями, но и с углеводно-фосфатной основой молекулы нуклеиновой кислоты. При этом происходит её денатурация. Полагают, что таким образом может взаимодействовать с нуклеиновыми кислотами в частности формальдегид.
2. Нарушение конформации нуклеиновых кислот.
Многие ксенобиотики образуют нековалентные связи с ДНК. При этом меняется конформация макромолекул. Так, известно высокое сродство к нуклеиновым кислотам производных акридина, которые, встраиваясь в молекулу ДНК между соседними парами оснований (интеркалация), изменяют её структуру. Таков же, вероятно, механизм действия этидиумбромида, профлавина и др.
Антрациклин, хлорахин, актиномицин и некоторые другие антибиотики также изменяют конформацию нуклеиновых кислот, не образуя с ними ковалентных связей.
3.3. Взаимодействие токсикантов с липидами
Важнейшая функция липидов - формирование биологических мембран. Вещества, разрушающие, изменяющие структуру липидов, нарушающие взаимодействие между молекулами липидов (гидрофобные связи) повреждают биологические мембраны и поэтому называются мембранотоксикантами. К числу таких относятся многие спирты, предельные и галогенированные углеводороды ("неэлектролиты"), детергенты (поверхностно-активные вещества), а также яды, обладающие фосфолипазной активностью (яды змей и т.д). Ряд токсикантов оказывает опосредованное мембранотоксическое действие, повышая уровень внутриклеточного Са2+, активируя эндогенные фосфолипазы, свободнорадикальные процессы в клетках (см. ниже) и т.д.
3.4. Взаимодействие токсикантов с селективными рецепторами
3.4.1. Селективные рецепторы клеточных мембран
Селективные рецепторы клеточных мембран - это протеины, встроенные в липидные бислои. Гидрофобный участок
- формирующие ионные каналы;
- связанные с G-протеинами;
- обладающие тирозинкиназной активностью;
- образующие межрецепторные сети.
1. Рецепторы формирующие ионные каналы. Эти рецепторы участвуют в передаче нервных импульсов в центральной нервной системе и на периферии. Обычно рецепторы данного типа состоят из нескольких субъединиц пронизывающих всю толщу биологической мембраны. Одна - две из этих субъединиц представляют собой собственно рецепторную область, связывающуюся с лигандом. Другие субъединицы после взаимодействия рецепторной области с лигандом изменяют свою конформацию и участвуют тем самым в формировании ионного канала. К числу известных каналобразующих рецепторов относятся никотинчувствительный рецептор ацетилхолина (Н-холинорецептор), ГАМК-ергический, глицинергический рецепторы. Первый из упомянутых является каналом для ионов Na+, два других - для ионов Cl-. Известно большое количество веществ, действующих на эти рецепторы. Например, курарин, никотин, анабазин (действуют на холинорецепторы), бициклофосфаты, норборнан, пикротоксинин (действуют на ГАМК-рецепторы), стрихнин (действует на рецепторы к глицину).
К этой же группе рецепторов можно отнести Na+-, К+, Ca2+- каналы возбудимых мембран, для которых пока не найдены эндогенные химические агонисты. Тем не менее, ионные каналы имеют рецепторную область связывания высокотоксичных ядов животного происхождения, таких как тетродотоксин, сакситоксин, батрахотоксин и др.
Первым из достаточно изученных каналобразующих рецепторов был никотинчувствительный рецептор ацетилхолина. Этот рецептор состоит из четырех типов субъединиц, из которых ацетилхолинсвязывающая
Клонирование и секвенирование генов, ответственных за синтез субъединиц никотинового рецептора, выделенного из различных органов животных одного вида и органов разных видов, позволили высказать предположение об эволюционной близости их происхождения. Об этом же свидетельствует и то обстоятельство, что не смотря на имеющиеся различия строения рецепторной области Н-холинорецепторов, выделенных из разных органов и тканей животных различных видов, их чувствительность к ацетилхолину, его агонистами и антагонистам различается несущественно.
2. Рецепторы, связанные с G-протеинами. Особый вариант передачи регуляторных сигналов представлен механизмом взаимодействия эндогенных лигандов с рецепторами, ассоциированными с G-протеинами (регуляторными протеинами). В этом случае сигналы, вызванные действием лиганда, приводят к конформационным изменениям рецепторного белка, затем переносятся на белки сопряжения, которые в свою очередь, уже или стимулирует или угнетает эффекторную систему в целом. Белки сопряжения в ходе передачи сигнала связывают молекулу гуанозинтрифосфата (ГТФ) и расщепляют её не гуанозиндифосфат и фосфат (отсюда название - G-протеины). Стимулирующие G-протеины (GS), активируют в ходе передачи сигнала аденилатциклазу клеток-эффекторов, а ингибиторные (GI) - угнетают этот энзим. Поскольку G-протеины расщепляют ГТФ, они называются также ГТФ-азами.
К числу рецепторов такого типа относятся в частности мускаринчувствительные холинэргические рецепторы (М-холинорецепторы),
Комплекс G-протеинов состоит из трех субъединиц. Наибольшая, с молекулярной массой 40000, называется
Функционирование системы передачи сигнала с помощью G-протеинов наилучшим образом изучено на примере
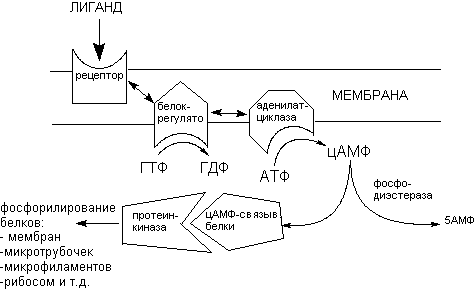
Рисунок 5. Схема проведения сигнала, через систему рецептора связанного с G-протеинами
В невозбужденном состоянии
Токсиканты могут модифицировать описанный процесс на любом из этапов проведения сигнала. Например, холерный и коклюшный токсины вызывают АДФ-рибозилирование GS-протеина после его связывания с молекулой ГТФ. В итоге развивается стойкая активация аденилатциклазы и перевозбуждение соответствующих клеток слизистых оболочек.
В настоящее время известно огромное количество веществ синтетических и естественного происхождения, избирательно взаимодействующих с рецепторами данного типа. Среди них многочисленные лекарственные средства (действующие на холинэргические, катехоламинергические, серотонинергические синапсы), интоксикация которыми развивается как при перевозбуждении, так и блокаде рецепторов (см. курс фармакологии). Сюда же относится большая группа веществ, обладающих психодислептической активностью (диэтиламид лизергиновой кислоты, псилоцин, псилоцибин, буфотенин, мескалин, хинуклидинилбензилат - BZ и т.д.).
3. Рецепторы с тирозинкиназной активностью. К рецепторам данного типа относятся, в частности, рецепторы к инсулину и гормону роста. Эти рецепторы состоят из одной или двух белковых субъединиц, закрепленных в липидном бислое с помощью пептидной
После взаимодействия с лигандом рецепторная молекула частично погружается в клетку. При этом активируется тирозинкиназная активность специальной субъединицы рецептора, либо энзима, тесно связанного с ним. Пусковым сигналом процессов, приводящих к активации клетки, является фосфорилирование внутриклеточных белков (по молекуле тирозина).
Токсиканты, избирательно действующие на данный тип рецепторов, пока не известны.
4. Рецепторы, образующие межрецепторные сети. В отличии от других мембраносвязанных рецепторов, связывание с лигандом в данном случае не приводит непосредственно к передаче сигнала на эффекторную систему. Процесс восприятия сигнала связан с образованием межрецепторных цепей на поверхности клеток. Наилучшим образом в группе рецепторов данного типа изучены рецепторы к Fc-фрагменту (fragment crystalline) антител (иммуноглобулинов). Эти рецепторы представляют собой агрегаты двух или более белковых субъединиц, молекулярная масса которых составляет около 30000 - 50000. Особое патофизиологическое значение имеет образование межрецепторных сетей IgE-Fc-рецепторов, поскольку это явление, наступающее вследствие связывания антигена с молекулами фиксированных на мембранах тучных клеток антител, приводит к взрывоподобному экзоцитозу гистамин-содержащих гранул. Экзоцитоз биологически активных веществ, и в частности, гистамина из тучных клеток - основа анафилактических реакций (см. ниже).
4. Изучение локализации рецепторов в биообъекте
Выяснение характера распределения рецепторов различных токсикантов в тканях, клетках, субклеточных структурах возможно с помощью различных методических приемов. Непрямым методом является обнаружение в исследуемом материале эндогенных биорегуляторных веществ, аналогами которых является токсикант, или ферментов их обмена. Как правило, для этой цели используют гистологические методы (гистохимия, иммуногистохимия). Такими способами, например, выявляют области синтеза, депонирования или разрушения нейромедиаторов (ГАМК, серотонина, дофамина, норадреналина, гистамина и т.д.). Однако на основании полученных результатов неправомочно делать заключение о том, что здесь же залегают и основные рецепторные образования к соответствующим токсикантам. Количество выявляемого медиатора в ткани (клетке) ни в коем случае не является характеристикой плотности рецепторов к исследуемому веществу.
Часто для выявления рецепторов в органах и тканях используют метод авторадиографии. При этом о характере их распределения часто судят по особенностям связывания токсикантов, меченых радиоактивными изотопами (радиолигандов). С помощью этого метода возможно изучение и внутриклеточной локализации рецепторов.
Метод компьютерной томографии, в частности позитронно-эмиссионная компьютерная томография, позволяет прижизненно изучать локализацию рецепторов токсикантов в тканях. Для этого в организм обследуемого вводят рецепторспецифичные радиолиганды (сам токсикант или его аналоги), меченные радионуклидами, испускающими позитроны (11С, 18F), локализацю которых в различных органах, по прошествии некоторого инкубационного периода, и выявляют с помощью соответствующей аппаратуры. С помощью такого подхода можно изучить распределение рецепторов в любом интересующем органе. Недостатками метода являются его недостаточно высокая разрешающая способность и дороговизна.
5. Понятие полирецепторного профиля связывания токсиканта
Селективное связывание токсиканта с рецепторами одного типа характерно лишь для очень небольшого числа высоко токсичных соединений (например, некоторые ФОС, ботулотоксин, сакситоксин, тетродотоксин, аманитин). Часто вещество имеет примерно одинаковое сродство к нескольким рецепторам, взаимодействие с которыми и приводит к формированию вполне определенного биологического эффекта (профиля токсических реакций). В этой связи, особенности проявлений интоксикации одним и тем же веществом, но различных степеней тяжести, обусловлено не только увеличением количества рецепторов одного типа, связавшихся с токсикантом, но и расширением спектра вступивших во взаимодействие рецепторов. По этой причине, часто, зная проявления интоксикации, мы не можем точно определить, каков механизм их формирования.
Сказанное выше относится к токсикантам с различными механизмами токсического действия, в том числе и влияющим на нейромедиаторный аппарат. В малых дозах эти ксенобиотики обычно избирательно взаимодействуют с рецепторами какого-то одного типа (например, нейролептики, трициклические антидепрессанты - ТАД - вмешиваются в передачу нервных импульсов в дофаминергических синапсах мозга), однако в высоких дозах действие распространяется и на другие синаптические структуры (упомянутые вещества обладают выраженной холинолитической активностью). В ряде случаев токсический эффект в большей степени связан с взаимодействием ксенобиотика с менее чувствительными, но более значимыми для поддержания гомеостаза рецепторами. Так, интоксикация упомянутыми нейролептиками и ТАД в основном проявляется эффектами, обусловленными блокадой холинэргических структур (психодислептические эффекты, вегетативные нарушения, устраняемые в значительной степени ингибиторами холинэстеразы).
В связи с изложенным, важным, при изучении механизмов токсического действия веществ, является определение рецепторного профиля их действия в широком диапазоне доз. В ходе этих исследований изучают виды рецепторов, с которыми может вступить во взаимодействие токсикант и количественные характеристики этого взаимодействия. Обычно задача решается с помощью радиолигандных методов исследования. Сравнение биологических эффектов, вызываемых действием на организм близких по строению, но различающихся по рецепторному профилю токсикантов, позволяет, с определенными оговорками, оценить значение каждого из механизмов формирования токсического процесса.
6. Радиолигандные методы изучения процесса взаимодействия токсиканта с рецепторами
Поскольку токсичность веществ во многом определяется их способностью взаимодействовать с рецепторами определенных типов, количественная оценка сродства конкретного вещества к конкретному рецептору порой имеет решающее значение для выявления механизма его токсического действия.
Количественно оценить сродство токсиканта к рецептору можно с помощью радиолигандных методов исследования. При этом, однако, эффекты, развивающиеся вследствие взаимодействия, остаются вне поля зрения исследователя. Более того, в ходе работы не представляется возможным решить вопрос, является ли исследуемое вещество агонистом или антагонистом (активатором или ингибитором) данного рецептора. Тем не менее, сочетание метода с биохимическими и физиологическими методиками позволяет получить развернутую картину механизма действия токсиканта и формирования ответной реакции биосистемы. К числу основных методов относятся: а) с полным насыщением рецептора и б) методы замещения радиолиганда.
а). Принцип метода с полным насыщением рецептора состоит в добавлении в инкубационную среду, содержащую рецептор, меченного изотопом токсиканта (радиолиганда) в возрастающей концентрации. Метод пригоден для исследования свойств веществ, прочно фиксирующихся на рецепторе (например, холинолитиков: скополамин, атропина, дитрана и т.д.). В ходе экспериментов с насыщением изучают зависимость количества образовавшегося радиолиганд-рецепторного комплекса (RL) от концентрации радиолиганда (L) при постоянном содержании в среде соответствующих рецепторов (R). После получения необходимых данных (как правило, представляемых в графической форме), можно рассчитать количественные характеристики процесса (константу диссоциации комплекса KD), используя общие положения закона действующих масс.
KD = [ R] [ L] /[ RL] (1)
Поскольку общая концентрация рецепторов, принимавших участие во взаимодействии, представляет собой сумму свободных и связавшихся рецепторов, т.е.
RO = [ R] + [ RL] (2)
преобразование уравнения (1) приводит к виду
[ RL] = [ RO] [ L] /(KD + [ L] ) (3)
Полученное уравнение, по сути, идентично уравнению Михаэлиса - Ментен, используемому при описании кинетики ферментативных процессов.
При концентрации свободного лиганда равной величине константы диссоциации комплекса лиганд-рецептор, имеем
[ RL] = [ RO] [ L] /[ L] + [ L] = 0,5[ RO] (4)
Таким образом, в упрощенной форме KD равна концентрации линганда, при которой половина рецепторов приняла участие в образовании лиганд-рецепторного комплекса.
Незначительные преобразования уравнения (3) приводят к линейной зависимости между исследуемыми показателями, что значительно упрощает анализ:
[ RL] /[ L] = -[ RL] /KD + [ RO] /KD (5)
Часто для обозначения количества лиганда, связавшегося с рецептором (RL), используют символ (В); свободного лиганда (L) - (F); общего количества рецепторов (RO) - (BMAX). Представленное в этих символах выражение (5) выглядит следующим образом:
B/F = -B/KD + BMAX/KD (6)
Исходя из этого выражения, можно рассчитать величину KD:
KD = (BMAX - B)F/B
Зависимость, построенная в координатах (BMAX - B) - по оси "Х", B/F - по оси "У", как правило, носит линейный характер. Угол наклона прямой позволяет определить величину KD:
B/F = 1/KD (BMAX - B)
На рисунке 6 представлены данные радиолигандного исследования связывания мощного холинолитика хинуклидинилбензилата (меченного тритием) с холинорецепторами сердечной мышцы свиньи. Анализ кривых позволяет определить сродство токсиканта к рецепторам соответствующего типа.
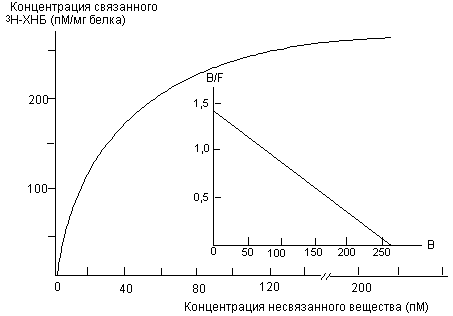
Рисунок 6. Кривая связывания радиолиганда [ 3Н] -хинуклидинилбензилата (3Н-ХНБ) на солюбилизированных мускаринчувствительных холинорецепторах сердца свиньи (G.S. Herron et al., 1982)
Уже в ходе таких простых исследований возникает проблема определения специфичности связывания лиганда. Неспецифическое связывание определяют путем внесения в среду 100 - 1000-кратного избытка не меченного изотопом вещества. Некоторые лиганды, особенно белковой природы, имеют свойство образовывать большое количество неспецифических связей с биосубстратом, иногда до 50% от общего количества образовавшихся комплексов.
б). Исследования методом радиолигандного замещения открывают путь к изучению процесса взаимодействия рецепторов с лигандами, характеризующимся формированием нестабильного комплекса. Это, прежде всего, характерно для веществ-агонистов соответствующих нейромедиаторов (например, холиномиметиков: никотина, ареколин, карбахола и т.д.). Принцип метода состоит в добавлении в инкубационную среду, содержащую рецептор и радиолиганд-агонист, немеченого изотопом вещества-антагониста в возрастающих концентрациях, до тех пор, пока лиганд полностью не будет вытеснен из связи с рецептором. Концентрация лиганда в опыте должна быть близка величине константы диссоциации комплекса лиганд-рецептор. В ходе эксперимента обычно определяют концентрацию антагониста, необходимую для вытеснения из связи 50% агониста рецептора. Эта характеристика обозначается как IC50. Константа диссоциации комплекса антагонист-рецептор (К1) и величина IC50 определяются величиной константы диссоциации агонист-рецепторного комплекса. Если предположить, что количество связанного радиолиганда состовляет лишь незначительную часть вещества в инкубационной среде, а также, что константа диссоциации комплекс значительно превышает величину ВMAX, то имеем:
К1 = IC50/(1 + F/KD)
Сравнивая величины констант диссоциации веществ, оценивают их сродство к изучаемому рецептору.
<< Содержание |